
Cultivons notre jardin
Pour créer des plantes virtuelles, les informaticiens travaillent à partir de travaux de botanique. Ainsi, ils peuvent simuler la croissance des plantes suivant leur emplacement, la lumière reçue… et habiller un paysage.
Pour les besoins de l’image de synthèse, les informaticiens se sont intéressés à la simulation en trois dimensions des plantes pour habiller les paysages virtuels dans lesquels la végétation manquait. Or l’architecture végétale est une affaire de botanique et les modèles architecturaux des arbres issus des travaux de Francis Hallé et Roelof Oldeman (1970) apportent les connaissances nécessaires aux informaticiens pour construire des plantes virtuelles, fidèles à la réalité. La construction 3D d’une plante se fait en couplant l’organogénèse à des algorithmes géométriques (translations, rotations) qui positionnent correctement les organes dans l’architecture, d’après des règles botaniques. Le volume et la forme des organes sont le résultat des relations sources-puits pendant la photosynthèse. La visualisation d’une plante utilise les outils classiques de rendu de l’image de synthèse (coloration, texturage, ombrage, réflexion…). Le laboratoire Botanique et bio-informatique de l’architecture des plantes (Amap) a développé une plateforme informatique qui permet de visualiser des paysages virtuels.
Au niveau cellulaire, la croissance d’une plante est assurée par deux mécanismes : la multiplication des cellules dans les « méristèmes » terminaux, qui sont contenus dans les bourgeons et qui fabriquent les organes (feuilles, entrenœuds, fruits, cernes, racines), et l’expansion de ces cellules qui permettent la croissance des organes. Au niveau de la plante, l’architecture est le résultat de ce double mécanisme : on parlera d’une part d’« organogénèse », pour caractériser la dynamique de la mise en place des nouveaux organes par les méristèmes, et de « croissance », pour caractériser l’expansion de ces organes jusqu’à leur maturation, grâce à l’action de la photosynthèse. On appelle encore « développement » la mise en place des nouveaux axes qui construisent l’architecture de la plante.
Les bases du bon jardinier
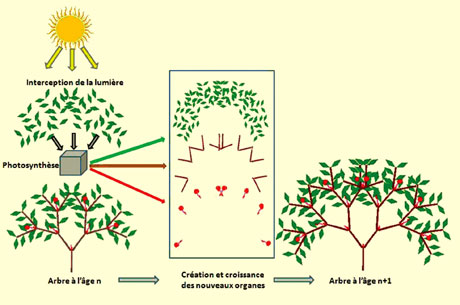
Croissance et architecture d’une plante (© Liama).
Une plante est constituée de 80 % d’eau, de 15 % de sucres et de 5 % d’oligo-éléments. C’est grâce à la photosynthèse qu’elle fabrique les sucres qui lui sont indispensables comme énergie et comme matériau pour sa croissance.
L’illustration présente le cycle de croissance d’un arbre : partant d’une architecture à l’étape n, les feuilles interceptent la lumière et produisent, par la photosynthèse, de la biomasse qui va alimenter l’organogénèse des bourgeons et assurer l’expansion des nouveaux organes. L’architecture de l’arbre à l’étape n + 1 est ainsi réalisée. Les modèles de l’agronomie postulent l’existence d’un pool commun de biomasse alimenté par la photosynthèse des feuilles (organes « sources »), qui distribue la matière végétale dans l’architecture pour assurer l’expansion des organes, feuilles, tiges, racines, cernes, fruits (organes « puits »). L’identification sur une plante des paramètres endogènes qui pilotent l’organogénèse et les relations sources-puits, ainsi que ceux qui calculent la photosynthèse (interception de la lumière par les feuilles et efficience de la lumière…), permet de simuler la croissance et l’architecture de la plante dans un environnement donné.
Au XIXe siècle, Nicolas Théodore de Saussure, chimiste et physiologiste suisse, a clarifié définitivement les problèmes liés à la nutrition tant en ce qui concerne le rôle des organes (feuilles, racines) que celui des constituants majeures de l’architecture végétale (sucres, eau, azote…). De même, Julius Sachs, botaniste allemand, a établi le fonctionnement respiratoire et photosynthétique des plantes. Alors que nombreux sont ceux qui pensent que les plantes n’ont besoin pour vivre que d’eau et d’air atmosphérique, il montre que la respiration s’accompagne de la destruction des sucres et que la synthèse de l’amidon provient de l’activité chlorophyllienne. Il établit l’équation fameuse qui lie la production de sucres avec les entrées de gaz carbonique dans les feuilles et de l’eau par les racines.
Développement et croissance ne peuvent exister que par l’approvisionnement du système plante en matière végétale. Une plante est constituée en moyenne de 80 % d’eau, provenant du sol et assimilée par les racines, et de 15 % de sucres (cellulose), composés d’eau et du gaz carbonique issu de l’atmosphère et assimilé par les feuilles. Le reste ou oligo-éléments, 5 % (azote, phosphore, potassium…), est indispensable aux mécanismes cellulaires, mais secondaire dans la constitution de la biomasse. La fabrication des sucres nécessite de l’énergie sous forme de lumière captée par les feuilles : la photosynthèse. Une particularité des plantes est que les sucres issus de la photosynthèse sont à la fois un carburant pour leur fonctionnement et un matériau pour leur construction sous forme de cellulose, dans des proportions qui peuvent varier. Suivant les conditions environnementales, la croissance peut être arrêtée. Par exemple, une plante en milieu salin consacre davantage son énergie à lutter contre l’invasion du sodium ; la synthèse de la cellulose est bloquée.
Les botanistes Francis Hallé et Roelof Oldeman ont réussi vers 1970 à étendre aux architectures des arbres le principe de la classification de la structure des inflorescences ; les fleurs sont disposées sur les tiges selon des motifs géométriques très réguliers. Il en est de même pour les branches des arbres qui s’empilent selon des modèles architecturaux qui dépendent du fonctionnement continu ou rythmique des bourgeons, de la floraison latérale ou terminale des branches et de leur type de différenciation que l’on appelle encore « âge physiologique ».
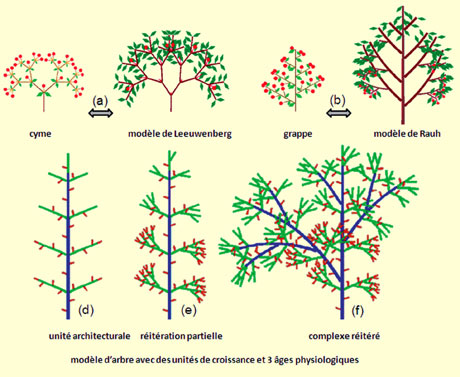
Analogie entre la structure des inflorescences et l’architecture des arbres (© Liama).
Les botanistes ont classé les arbres de toutes les espèces connues selon leur structure. Trente catégories de « modèles architecturaux » ont ainsi été créées. Ils permettent de prédire la manière dont va se développer tel ou tel arbre.
Les processus de croissance créent d’une façon combinatoire des « modèles architecturaux » dont le nombre n’excède pas trente. Ainsi la cyme de la petite centaurée a la même structure modulaire que celle d’un frangipanier (a) (ramification sympodiale). De même, à la grappe de fleurs d’un lilas on peut faire correspondre l’architecture d’un merisier (b) (ramification monopodiale). Mais les modèles ne sont que des schémas et il faut recourir à la notion d’« unité architecturale » (d) pour décrire plus précisément le schéma de développement des arbres. Les axes sont caractérisés par des « âges physiologiques » qui déterminent leur morphologie et sont constitués d’unités de croissance. La réitération des axes peut être partielle (e), sur les branches, ou totale (f), si le tronc est impliqué. Les bourgeons axillaires sont plus âgés physiologiquement que le bourgeon principal (sauf cas de réitération). Leur mutation au cours du fonctionnement en un âge physiologique supérieur produit le vieillissement de la structure.
Des racines aux branches, toute une architecture
Un même type d’axe peut se retrouver à différents endroits au sein d’une architecture. Ainsi sont nés les « modèles architecturaux », dessins de botanistes qui classent en moins de trente catégories les arbres de toutes les espèces connues. Ces modèles sont l’expression stable d’un programme génétique de développement. L’architecture végétale est ici considérée non pas d’une manière statique à l’instar des planches botaniques qui décrivent seulement minutieusement les parties de la plante, mais comme le résultat de la cinétique de la mise en place des axes feuillés d’un arbre par les méristèmes.
La modélisation de la production végétale a pour but de prédire les récoltes d’un point de vue quantitatif et d’optimiser les itinéraires culturaux. Initiée par le Hollandais de Witt (1970), elle s’appuie sur des paramètres climatiques (lumière, eau, température) et des indices végétaux, comme l’indice foliaire (surface des feuilles par m2) et l’indice de récolte (pourcentage de la biomasse utile sur la biomasse totale produite), pour mettre en équation la photosynthèse dont le résultat est une production de matière sèche par jour et par m2. Seule la production végétale au m2 est considérée et les plantes ne sont pas individualisées. À l’échelle de la plante, sans descendre au niveau cellulaire, et en ignorant les mécanismes physiologiques complexes de la photosynthèse et de la respiration, il se dégage des lois qui permettent de modéliser le comportement macroscopique du fonctionnement végétal.
Bien calculer sa récolte
Une loi empirique remarquable à l’échelle de la plante est la notion de « temps thermique ». Il correspond à la somme des températures moyennes journalières reçues par la plante. Le développement est très instable si on considère le temps calendaire, mais tout devient linéaire avec le temps thermique. Il faut un certain cumul de températures pour déclencher, au-delà d’un seuil, la levée des semences, la mise à fleur, l’émission des nouvelles feuilles par les méristèmes, etc. Le temps thermique pilote les séquences du développement végétal et il remplace souvent le temps calendaire dans les modèles de l’agronomie.
L’autre lien à étudier dans la production de matière sèche (MS) est celui entre la croissance et l’interception de la lumière. Sous sa forme la plus simple, cette équation (1) s’écrit MS = LUE.PAR.(1 – e– k LAI).
LUE est l’efficience de la lumière en conversion de matière sèche pour la partie active de la radiation (PAR) reçue par le couvert. Le LAI ou indice foliaire représente la surface foliaire par m2 mise en place par la culture. Plus cet indice est élevé, plus il y a de lumière interceptée par le feuillage. Le coefficient k est déterminé empiriquement. Le terme exponentiel modélise le recouvrement des feuilles les unes sur les autres qui diminue l’efficacité de la lumière.
Ce formalisme qui calcule la lumière interceptée par le feuillage correspond à la loi de Beer-Lambert. Au-delà de LAI = 4, toute la lumière est interceptée et la production en matière sèche d’un m2 de culture sature. Les feuilles supplémentaires ne sont plus efficaces et augmenter la densité de plantation ne sert à rien. On ne peut guère produire plus de 20 grammes de matière sèche par jour et par m2 dans des conditions favorables.
Mettre en lumière la croissance
Des simplifications judicieuses, démontrées par des expérimentations agronomiques, montrent que la prise en compte de l’architecture de la plante n’est généralement pas nécessaire. On postule l’existence d’un pool commun de biomasse. De la même façon, on ne considère que la photosynthèse nette, c’est-à-dire la proportion de sucres qui entrent comme matériaux de construction dans la matière sèche des organes. D’une façon générale, la production de matière sèche est divisée en compartiments (feuilles, tiges, racines, fruits) dont les proportions sont des « puits » pour le stockage de la matière produite. L’indice de récolte est la proportion utile de la production de matière sèche qui va au compartiment végétal d’un intérêt économique. L’équation (1) est la base des modèles de prédiction de récolte. En effet, elle cumule jour après jour la transformation en matière sèche de l’énergie reçue par le couvert.
Par ailleurs, les modèles structures fonctions des plantes proposés par l’informatique et l’agronomie intègrent l’architecture comme support du fonctionnement.
Ici, ce ne sont plus les compartiments végétaux qui sont visés, mais les organes individuels. Ainsi les branches sont constituées de « phytomères », entités élémentaires faites d’un entrenœud, d’une ou plusieurs feuilles et de bourgeons axillaires qui donnent naissance à des fruits ou des rameaux. Certains organes produisent de la matière, comme la feuille et la graine, ce sont des « sources », d’autres en consomment, les « puits », comme les entrenœuds et les fruits. Le développement de l’architecture est contrôlé par une grammaire qui pilote l’organogénèse en fonction du temps.
Créer des plantes modèles
La modélisation du fonctionnement végétal s’appuie sur un maillage qui permet une représentation géométrique de la plante avec des polygones (ceux des feuilles vont intercepter la lumière), ou sous une autre forme, avec des « éléments finis » pour assurer la propagation des sucres et de l’eau dans la structure. Les organes sources envoient de la matière dans le réseau et les organes puits interceptent cette matière selon leur « force de puits ». Il s’agit ici d’imiter le processus de morphogénèse dans son intégralité. Ces systèmes permettent de mieux connaître la plante, mais pas d’aborder le calcul d’une production à l’échelle agronomique. En effet, les temps de simulation deviennent trop conséquents, ce qui, ajouté à un nombre important de paramètres à identifier, ne permet pas commodément de calibrer le modèle en conditions réelles.
De la plante au champ
Un bon point de vue est d’utiliser les méthodes des modèles de production au niveau du m2 pour calculer la photosynthèse et de recourir aux modèles architecturaux pour la répartition de la matière dans les organes, au niveau des plantes individualisées. C’est ce qui est fait avec le modèle GreenLab. On prend en compte les simplifications établies par les agronomes dans le fonctionnement (pool commun, photosynthèse nette, loi de Beer…) ce qui permet de court-circuiter la structure topologique. La formule (1) est adaptée à l’échelle d’une plante et donne l’équation suivante (2) :
Q = LUE.PAR.Sp.(1 – e–k Sf/Sp)
Q est l’accroissement en biomasse de la plante par cycle, Sf est la surface foliaire de la plante et Sp un paramètre qui a la dimension d’une surface et qui permet à l’équation (2) de continuer à fonctionner même quand le couvert végétal n’est pas homogène. Pour une haute densité d de peuplement, le couvert est homogène et on montre que Sp = 1/d. Ceci est vérifié expérimentalement sur les grandes cultures. Les formules (1) et (2) sont alors équivalentes, mais la formule (2) permet le passage de la plante au peuplement.
L’organogénèse est pilotée par les équations de production des méristèmes qui sont déduites des observations. Croissance, mortalité, ramification, sont décrites au moyen de probabilités. À chaque cycle de croissance, de nouvelles populations d’organes (cohortes) sont mises en place. Chaque organe prélève de la biomasse dans le pool commun selon une fonction force de puits qui varie pendant sa maturation. Tous les organes d’une même cohorte et d’un même type sont dans le même état, ce qui permet une factorisation très efficace de la demande D de la plante ou somme de tous les puits.
L’accroissement en biomasse ∆q d’un organe dépend de son puits, de l’offre Q calculée par l’équation (2) et la demande D selon la formule :
∆q = p.Q/D
Par sommation des accroissements, on obtient le poids de l’organe et par des règles géométriques, ses dimensions (par exemple la surface pour une feuille). Le poids des compartiments s’obtient en sommant les cohortes d’organes de même type. La surface foliaire de la plante Sf s’en déduit en particulier, ce qui permet à l’équation (2) de fonctionner.
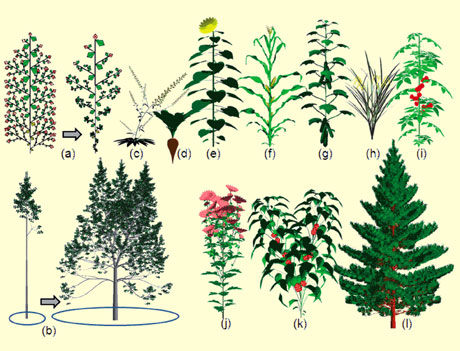
Simulation de la croissance des plantes et de leur plasticité avec le modèle GreenLab (© Digiplante – Liama).
Ces plantes virtuelles sont obtenues grâce à un logiciel qui tient particulièrement compte de la manière dont les organes (feuilles, racines, fruits…) évoluent, ce qui peut être très différent suivant les conditions environnementales.
Le modèle GreenLab intègre les connaissances botaniques et agronomiques nécessaires et suffisantes à la simulation de l’architecture des plantes. C’est un des rares modèles qui travaille au niveau de l’organe et qui est basé sur des données expérimentales. Chez les herbacées, les schémas de développement sont très réguliers, mais la croissance des organes ne suit pas l’ordre de leur création (a). Chez les arbres, le schéma de développement peut être très perturbé et sous la dépendance de la photosynthèse : un hêtre de 15 ans peut faire 15 cm ou 15 m selon qu’il est à l’ombre ou au soleil ! La densité de plantation en général ne joue pas sur la hauteur de l’arbre, mais sur la durée de vie des branches et la réitération (b). Ce modèle est utilisé dans plusieurs centres de recherche sur de nombreuses plantes cultivées : (c) Arabidopsis, (d) betterave, (e) tournesol, (f) maïs, (g) concombre, (h) riz, (i) tomate, (j) chrysanthème, (k) poivron, (l) pin.
Il s’agit donc d’un modèle dynamique de croissance des plantes qui fonctionne par rétroaction, entre la croissance et le développement. Le calcul de la production de la plante n’a pas besoin de s’appuyer sur le détail de l’architecture, mais seulement sur les équations de productions et les relations sources-puits, ce qui fait gagner beaucoup de temps de calcul et permet de mettre en œuvre les méthodes de calibration du modèle, l’optimisation et le contrôle.
La modélisation de la croissance des plantes offre des perspectives intéressantes aux sciences de l’environnement. Pour l’agriculture, il s’agit de fournir de nouveaux paramètres à la génétique, pour caractériser plus finement le fonctionnement des espèces cultivées et orienter les croisements. Ces mêmes paramètres sont utilisables dans les équations dynamiques du modèle pour optimiser les itinéraires culturaux (irrigation, traitements). Enfin, pour le paysagisme et l’urbanisme, on peut concevoir des bibliothèques de plantes sensibles à l’environnement naturel qui pourraient être pilotées par des systèmes d’information géographique (SIG). Les « paysages virtuels » créés par la synthèse d’image se transformeraient alors en « paysages fonctionnels » ouverts aux différents scénarios climatiques.
Livres
- Hallé F. (ss dir.), « Modélisation et simulation de l’architecture et de la production végétale », in Aux origines des plantes, Fayard, 2008.
- Varenne F., Du modèle à la simulation informatique, coll. « Mathesis », Vrin, 2007.
Sites web
- Amap, botanique et bio-informatique de l’architecture de plantes
- Équipe Digiplante, architecture et production végétale
Cet article est paru dans la revue DocSciences n°8 Le numérique et les sciences du vivant, éditée par le CRDP de l’Académie de Versailles en partenariat avec l’Inria.
Newsletter
Le responsable de ce traitement est Inria. En saisissant votre adresse mail, vous consentez à recevoir chaque mois une sélection d'articles et à ce que vos données soient collectées et stockées comme décrit dans notre politique de confidentialité
Niveau de lecture
Aidez-nous à évaluer le niveau de lecture de ce document.
Votre choix a été pris en compte. Merci d'avoir estimé le niveau de ce document !

Philippe de Reffye