
Modéliser le vivant : créer des plantes virtuelles pour comprendre, simuler, tester
Avant d’entrer dans le vif du sujet, et aborder les caractéristiques des plantes virtuelles, il convient de donner quelques repères sur les mécanismes biologiques étudiés grâce à ces modèles.
1. Une plante : comment ça pousse ?
La croissance des plantes en bref
À l’échelle des axes, la croissance est le résultat de deux processus : d’une part de nouvelles cellules et organes sont formés ; d’autre part ces cellules s’allongent et se spécialisent en fonction de leur position.
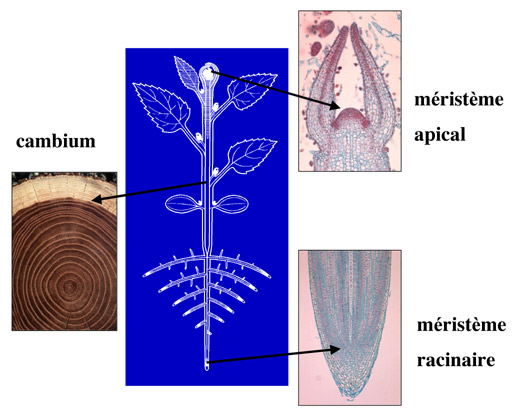
Les méristèmes. Les méristèmes apicaux sont situés aux extrémités – apex – des tiges et des racines. Le cambium est un méristème secondaire qui permet à la tige de croître en diamètre.
Chez les plantes, les divisions cellulaires se produisent dans des territoires particuliers, situés aux extrémités des tiges et des racines et appelés méristèmes. Dans les méristèmes, les divisions se déroulent tout au long de la vie des plantes et, contrairement aux animaux dont la faculté à produire de nouveaux organes s’achève avec la phase embryonnaire, les plantes conservent leur capacité à produire de nouvelles feuilles, portions de tiges et fleurs pendant plusieurs mois voire plusieurs années. Les feuilles sont formées régulièrement autour du centre du méristème, qui reste quiescent (c’est-à-dire inactif) tant que la floraison ne se produit pas. Lorsque cette étape clé de la production se déclenche, les divisions envahissent le centre du méristème qui donne alors naissance successivement aux sépales, pétales, étamines, pistil et ovaire. La floraison marque alors la fin de la capacité d’organogenèse de chaque méristème aérien. Mais quelle belle fin de se transformer en fleur odorante et colorée, rose ou coquelicot ou encore orchidée ! À moins que le méristème ne se transforme en une vrille élégante ? Chez les végétaux bien des solutions sont possibles…
Les divisions cellulaires aux extrémités des axes permettent aux plantes de s’élever vers la lumière et ceci en dépit de la gravité.
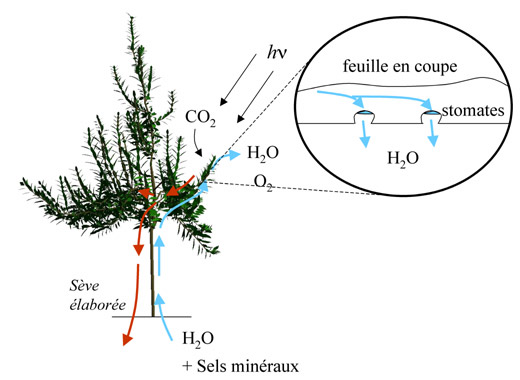
Principe du transport de l’eau et de la sève élaborée.
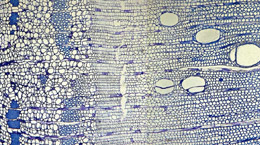
Détails du phloème (à gauche) et du xylème (à droite).
Pour cela, les plantes disposent d’un second tissu méristématique, lui aussi capable de divisions, qui produit les tissus assurant à la fois le soutien des tiges et la conduction de l’eau et des aliments nutritifs entre les organes aériens et souterrains. Ce méristème, dit secondaire, et appelé aussi cambium, est disposé en manchon le long des axes et permet la croissance en épaisseur. Dans ce tissu, les divisions se produisent de façon polarisée c’est à dire d’une part vers l’intérieur de l’axe et d’autre part vers l’extérieur, sous l’écorce. Vers l’intérieur des axes, les cellules méristématiques donnent naissance aux vaisseaux conducteurs du xylème qui permettent le transit de l’eau depuis les racines vers les feuilles et les fruits. Le xylème est constitué de cellules mortes et imprégnées de lignine, substance rigidifiant les parois cellulaires et améliorant à la fois leur capacité de soutien et de transfert de l’eau. Ainsi, les cellules du xylème se comportent comme des tuyaux dans lesquels l’eau est aspirée passivement par la transpiration des feuilles. Vers l’extérieur de l’axe, le méristème secondaire donne naissance aux tissus conducteurs du phloème. Les cellules de ce tissu vivant transportent activement la sève élaborée, amenant les sucres issus des feuilles vers les autres organes.
À ces deux processus de croissance, primaire et secondaire, s’ajoute le plus souvent chez les plantes, la capacité à se ramifier. La ramification correspond à l’entrée en croissance des territoires méristématiques dits axillaires car disposés à l’aisselle des feuilles. La disposition des rameaux axillaires est elle aussi variable et conduit à des formes arborescentes diverses. Les plantes ayant tendance à produire les rameaux les plus longs vers la base des rameaux porteurs ont généralement des formes buissonnantes, alors qu’une disposition des rameaux les plus longs vers le sommet du rameau porteur donnera des arbres de grandes dimensions.
Petites plantes ou arbres ?
Les agronomes distinguent les plantes annuelles, qui poussent sur une période courte (typiquement en moins d’une année) et les plantes pérennes, dont la durée de vie se poursuit sur plusieurs cycles de croissance et peut atteindre des valeurs bien supérieures à la durée de vie humaine. Cette variation de la durée de vie des végétaux résulte de leur capacité à maintenir les deux types de croissance, primaire et secondaire, sur une période plus ou moins longue. Ainsi, l’arabette, Arabidopsis thaliana, plante choisie comme « modèle » par la communauté scientifique, est composée seulement d’une rosette de feuilles et d’une tige à floraison terminale. Son cycle s’effectue en quelques mois. D’autres plantes, de dimensions presque aussi discrètes que l’arabette, sont pourtant des plantes pérennes, comme par exemple la luzerne de nos prairies. Enfin, lorsque la croissance en hauteur, la croissance secondaire et la ramification s’allient, l’organisme peut atteindre des dimensions considérables. Ce dernier cas concerne la plupart des arbres dont les cimes majestueuses contribuent au plaisir de nos promenades dans les parcs naturels ou aménagés.
Les plantes pérennes présentent également la capacité à survivre pendant les périodes climatiques défavorables. En régions de climat tempéré, elles devront supporter la période hivernale. Dans d’autres régions, elles devront par exemple supporter de longues périodes de sécheresse. La capacité de survie est généralement associée à la capacité à interrompre les cycles de division cellulaire et d’allongement, et à entrer dans un état dit dormant selon son degré d’interaction avec l’environnement. En outre, de nombreuses plantes ont développé des organes particuliers dont la fonction est de protéger les zones méristématiques. Ainsi, les bourgeons sont-ils constitués d’écailles, qui sont des feuilles modifiées qui, en emprisonnant de l’air autour des jeunes ébauches foliaires et du méristème, les protègent contre le froid. La pérennité des structures végétales sous des climats fluctuants est donc souvent le résultat d’une croissance dite rythmique car présentant alternativement des phases de croissance et de repos.
Au cœur de la croissance, les méristèmes
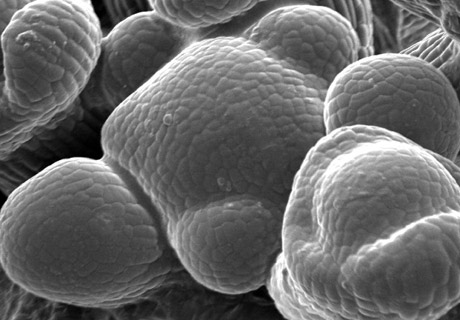
Méristème d’inflorescence d’Arabidopsis thaliana.
Le méristème d’inflorescence a une structure en forme de dôme. Les organes sont produits sur les flancs de la structure méristématique selon un schéma spatial précis, appelé phyllotaxie. Dans le cas du méristème d’inflorescence d’Arabidopsis thaliana, il s’agit d’une phyllotaxie spiralée.
Photo prise en microscopie électronique à balayage.
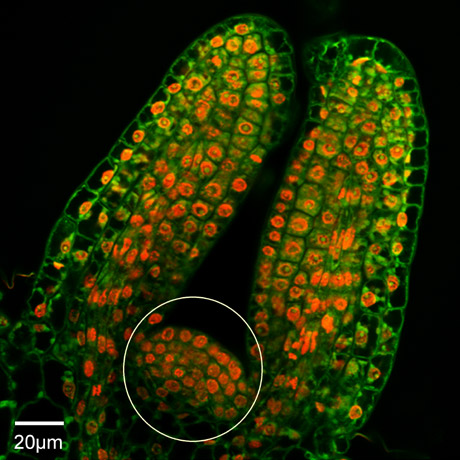
Coupe longitudinale qui traverse un jeune apex.
Le méristème apical (encerclé) a déjà généré deux jeunes feuilles. Les noyaux sont colorés en rouge, le cytoplasme des cellules en vert.
L’élucidation des mécanismes contrôlant le fonctionnement méristématique est depuis longtemps un enjeu majeur de la biologie du développement des plantes. Nous nous intéresserons ici à un méristème particulier, appelé le méristème apical caulinaire, qui initie toute la partie aérienne de la plante.
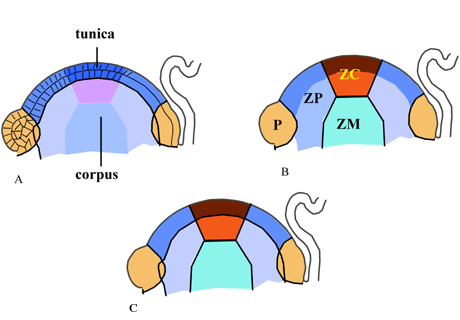
Modèle d’organisation du méristème apical caulinaire des angiospermes.
L’exemple illustré ici est celui des dicotylédones.
(A) modèle d’organisation en assise. La tunica est composée de 2 assises L1 et L2. Le corpus est également appelé L3.
(B) modèle d’organisation en zones concentriques. ZC : zone centrale ; ZP : zone périphérique ; ZM : zone médullaire ; P : primordium.
(C) superposition des deux modèles.
Tous les méristèmes apicaux caulinaires des végétaux supérieurs sont pluricellulaires. Cependant, au sein du méristème, les cellules qui constituent les unités de base construisant le méristème ne fonctionnent pas de façon autonome. Elles sont organisées en domaines multicellulaires permettant le fonctionnement du méristème comme un ensemble coordonné. Ces domaines distincts au niveau cytologique représentent des unités fonctionnelles.
Des approches génétiques, menées au cours de la dernière décennie, principalement chez l’espèce modèle Arabidopsis thaliana mais aussi chez le maïs, ont révélé une partie des mécanismes moléculaires qui sous-tendent l’organisation du méristème apical caulinaire. Associées aux approches biochimiques et cellulaires, ces analyses génétiques ont établi une première base de la compréhension de son fonctionnement. Ainsi des gènes qui contrôlent la maintenance du méristème et l’initiation des primordia (c’est-à-dire des très jeunes organes) ont été identifiés. Un certain nombre de ces gènes est uniquement actif dans certaines cellules, et chaque domaine du méristème est caractérisé par une combinaison unique d’expression génique.
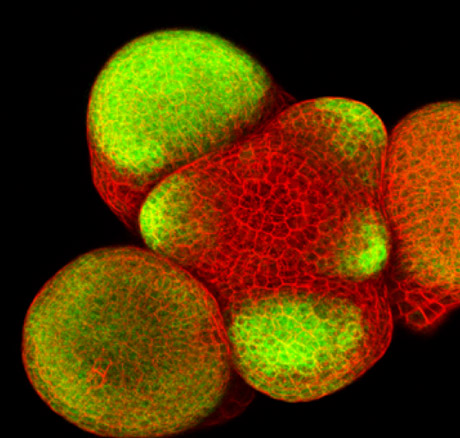
Expression de la protéine fluorescente GFP sous contrôle d’un promoteur méristématique actif dans les très jeunes primordia du méristème d’inflorescence d’Arabidopsis thaliana.
Apex d’une inflorescence sauvage coloré au iodure de propidium pour visualiser le contour des cellules. La GFP est exprimée spécifiquement dans les primordia, selon un profil spatial correspondant à celui du gène AINTEGUMENTA. Ce gène joue un rôle important dans la croissance des jeunes organes.
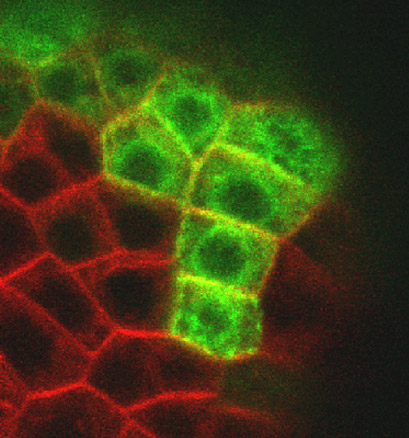
Détail montrant des cellules qui expriment et qui n’expriment pas le marqueur. Ceci montre que des cellules adjacentes peuvent se comporter très différemment.
Cette organisation très ordonnée se base sur des interactions constantes entre les cellules sous forme d’échanges de métabolites, de signaux hormonaux, ou de contraintes physiques. La complexité de ces réseaux d’interaction est telle que des approches de modélisation sont devenues indispensables afin de comprendre les mécanismes qui sont en jeu.
2. Des plantes virtuelles : qu’est-ce que c’est ?
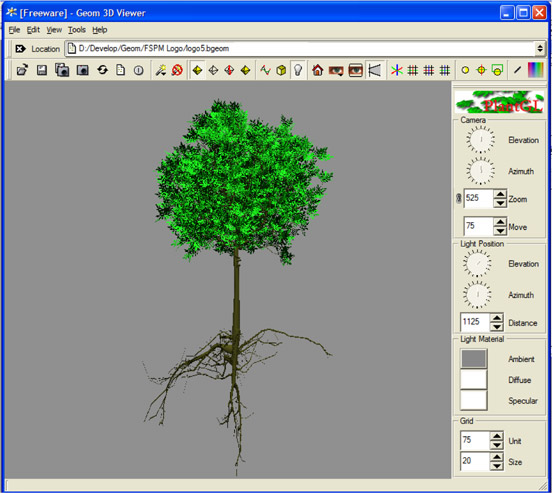
Des plantes virtuelles pour étudier, comprendre et réaliser des expérimentations virtuelles.
Une plante virtuelle est un objet informatique qui décrit la plante comme un ensemble d’organes, représentés par des formes géométriques placées dans l’espace, et connectés entre eux. Cette description informatique peut éventuellement intégrer l’évolution dans le temps de ces organes, et représenter alors une plante en croissance.
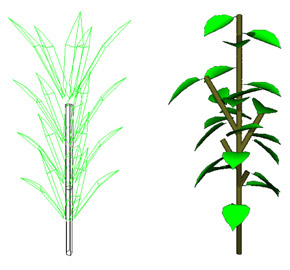
Utilisation de formes géométriques simples pour la représentation des organes.
Les organes choisis pour constituer la plante virtuelle sont le plus souvent de nature botanique : des entrenœuds, des feuilles, des unités de croissance, des branches, des tiges ou des troncs. Mais ils peuvent aussi être définis sans référence botanique : des segments de branche.
Les formes géométriques utilisées pour représenter un organe peuvent être simples : un polygone pour une feuille, un cylindre pour un tronc, un tronc de cône pour un segment de branche.
Mais elles peuvent être aussi définies avec des objets géométriques plus sophistiqués : des branches incurvées représentées par des courbes splines, définies par des points de contrôle et extrudées pour tenir compte de l’épaisseur de la branche.
Ces formes géométriques 2D ou 3D sont disposées dans l’espace 3D occupé par la plante, en tenant compte de la dimension des organes, de leurs coordonnées spatiales et de leur orientation angulaire (par les angles de rotation autour des 3 axes définissant le repère d’un espace 3D).
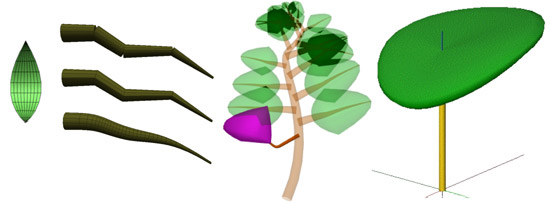
Utilisation de formes géométriques plus sophistiquées.
Les connexions physiques des organes les uns avec les autres – ce qu’on appelle la topologie de la plante – sont aussi prises en compte.
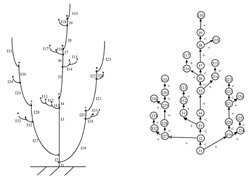
Représentation de la topologie d’une plante virtuelle.
Ces liaisons topologiques traduisent que tout constituant de la plante a été créé à partir du tissu d’un autre constituant, appelé parent. Si le tissu parental est un bourgeon terminal – situé à l’extrémité de l’organe parent – alors la connexion est une relation de succession : la succession des entrenœuds forme une branche. Si le tissu parental est un bourgeon axillaire – situé à la base du pétiole de toute feuille – alors la connexion est une relation de ramification. Succession et ramification sont les seules relations topologiques qui existent entre organes végétaux. Ces relations sont représentées par un graphe arborescent.
C’est à partir de cette représentation de la topologie que peut être étudiée la croissance des structures ramifiées.
Enfin, les constituants de l’arbre peuvent être définis à plusieurs échelles spatiales : du plus simple : un arbre décrit par un cylindre pour le tronc, surmonté d’une sphère pour la couronne ; au plus détaillé : la description de tous les segments de branche – y compris le tronc – et des feuilles constituant l’arbre. Les différentes échelles de description peuvent être intégrées dans une plante virtuelle unique, dite multi-échelles.

Trois échelles spatiales pour représenter la même plante.

Modèle photo-réaliste. P. Prusinkiewicz et al., 1997
Les plantes virtuelles peuvent être construites soit à partir de mesures de plantes réelles, soit à partir de modèles par simulation.
Elles permettent de décrire la complexité de la structure des plantes, d’une façon relativement simple, exhaustive et explicite. En plus des informations géométriques et topologiques, d’autres propriétés peuvent être attribuées aux organes : propriétés physiques, comme la texture de l’organe permettant un rendu réaliste pour des applications infographiques ; propriétés biologiques, comme les réponses physiologiques des organes aux signaux de l’environnement (sécheresse de l’air ou du sol, lumière, température…), ou encore la composition biochimique des organes.

Simulation de l’interception de la lumière par radiosité hiérarchique.
233 arbres, 2 millions de feuilles et 5 millions de polygones initiaux, ont nécessité 4 heures de calcul. Une approximation acceptable de l’illumination globale est obtenue en 20 mn.
C. Soler, F. Sillion et al. 2003
Une telle représentation informatique permet :
- de visualiser la structure 3D des plantes pour des applications infographiques ou biologiques ;
- de simuler des processus biologiques, représentés par des lois physiques, comme l’interception de la lumière en relation avec la distribution du feuillage dans l’espace 3D, ou par des lois biophysiques ou physiologiques, comme la circulation de l’eau ou des nutriments dans la plante, en relation avec sa topologie ;
- de représenter la croissance et le développement de la plante, en agissant sur les propriétés géométriques des organes et en ajoutant de nouveaux organes.
Tous les calculs sont possibles ; mais reste à connaître et pouvoir représenter sous forme mathématique les mécanismes biologiques déterminants.

Simulation du développement de l’architecture d’un arbre.
Y. Caraglio et J.F. Barczi, 2000.
Quelles sont les difficultés ?
Comme dans le cas de nombre de systèmes biologiques, la simulation numérique des plantes n’est pas aussi bien maîtrisée que celle des objets manufacturés (comme par exemple la simulation des efforts mécaniques sur un avion en vol).
Il n’est pas (encore ?) possible par exemple de prévoir le développement exact d’une plante, sa hauteur, son diamètre, la taille et l’orientation de chacune de ses feuilles, la position et le nombre de ses fleurs, à partir d’une graine donnée, dans des conditions de culture (température, humidité, lumière, éléments nutritifs) données. Ceci pour quatre raisons essentielles :
- les phénomènes biologiques impliqués dans le processus de développement sont très nombreux et leurs lois sont beaucoup moins bien connues que les lois élémentaires de la physique par exemple ;
- les niveaux d’organisation sont nombreux et étroitement enchevêtrés, rendant extrêmement difficile, voire arbitraire, la décomposition d’un tel système en sous-systèmes élémentaires plus simples et indépendants ;
- l’interaction avec l’environnement est complexe (sol, lumière, gaz, nutriments, insectes…) et module la croissance de la plante de manière continue, tout au long de sa vie ;
- les conditions initiales, comme l’état de la graine, de la plante réelle ou d’une cellule par exemple, ne sont pas mesurables à une précision arbitraire.
Ces raisons nous empêchent (peut être à jamais) de prédire « la trajectoire d’une plante » au cours de son développement de façon aussi précise que celle d’une fusée ou d’un avion. Pourtant, la construction de plantes virtuelles ouvre des perspectives nouvelles de compréhension et de gestion du vivant. Il devient par exemple possible :
- d’étudier l’interaction entre forme et processus biophysiques (analyse des flux internes à la plante par exemple ou des échanges avec l’environnement) sur les bases d’une représentation réaliste détaillée ;
- d’analyser et de mieux comprendre le couplage des différents phénomènes biologiques mis en jeu dans le fonctionnement ou la croissance (flux d’eau, photosynthèse, flux de sucres qui conduisent par exemple au stockage du carbone dans une plante, échange d’informations via des flux hormonaux entre différentes parties de la plante au cours de la croissance) ;
- de comparer sur des bases structurelles et quantitatives la structure ramifiée de plusieurs plantes ;
- de mieux prendre en compte le côté non-déterministe des mécanismes de croissance, en identifiant les différentes sources de bruit ou de non-connaissance des données à l’aide de modèles stochastiques appropriés ;
- de prendre en compte la forme de façon détaillée dans la relation entre l’expression phénotypique d’une plante et son matériel génétique (ou génotype).
3. Des plantes virtuelles : pour quoi faire ?
Demande de la société et maturité des technologies
La compréhension et le contrôle de la croissance des plantes ont toujours constitué des préoccupations centrales de l’activité humaine, pour se nourrir, pour se chauffer, pour construire, pour soigner, pour s’habiller, etc. Depuis longtemps, nombre de questions de société reposent ainsi sur l’amélioration des techniques de gestion et de prédiction de l’état, de la production ou de la croissance de la végétation. Au-delà des approches agronomiques traditionnelles, qui tentent d’optimiser de la production de biomasse (fruits, bois…), l’intérêt récent de la société pour les questions concernant l’écologie, le développement durable et le changement climatique appelle le développement d’une meilleure compréhension du rôle essentiel des végétaux dans ces questions. Les problèmes scientifiques associés sont nombreux, complexes et demandent de développer une nouvelle approche scientifique, pluridisciplinaire, dans laquelle il devra être possible de contrôler non seulement la production des plantes mais également la façon dont celle-ci est élaborée par les plantes au cours de leur croissance.
Pour étudier ces questions, les chercheurs développent des modèles du fonctionnement et de la croissance des plantes et les outils de simulation associés. Au cours de la dernière décennie, ce travail de recherche a été stimulé par le développement de la puissance des ordinateurs qui a permis aux technologies de modélisation tridimensionnelle (3D) d’atteindre une phase de maturité. Ceci, à son tour, a favorisé le développement de modèles informatiques 3D de plantes appelés « plantes virtuelles ». L’enjeu de ces modèles consiste, à travers l’utilisation de représentations 3D, à expliciter le rôle de la spatialisation des processus physiques et biologiques dans le fonctionnement et la croissance des plantes.
Aujourd’hui, où en sommes nous ?
Les techniques de développement de plantes virtuelles sont encore dans l’enfance. Elles progressent toutefois très rapidement et participent plus généralement à l’émergence d’une bioinformatique au sens large. Pour fixer les idées, nous savons aujourd’hui par exemple :
- mesurer et reconstruire des plantes en 3D dans un ordinateur de façon détaillée, organe par organe, par méthode de « pointage 3D » ;



Un arbre et sa reconstruction 3D obtenue à partir de mesures effectuées par pointage 3D.
- construire des modèles biophysiques précis (modèles d’interception lumineuse, et de mécanique) ;
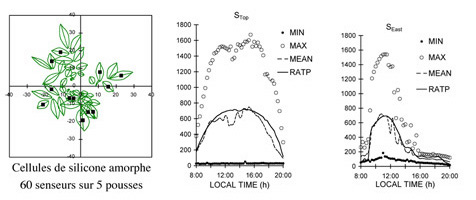
Évaluation d’un modèle d’interception. RATP : radiation absorption.
- construire de premiers modèles simulant quantitativement la photosynthèse, la circulation de l’eau et des sucres dans la plante, sur la base de règles de fonctionnement imparfaitement connues ;
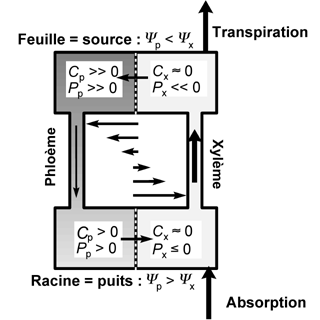
Modélisation du couplage entre les flux d’eau et de sucres à travers les différents tissus de la plante.
Le modèle bio-physique de Münch permet de modéliser ce couplage (C concentration en sucres, P pression de turgescence, Ψ potentiel hydrique). L’approche permet de simuler de façon mécaniste la circulation de flux au sein d’un système ramifié.
D’après Daudet et al., 2002 - simuler de façon très réaliste le développement qualitatif ou quantitatif d’une plante et le visualiser par infographie.
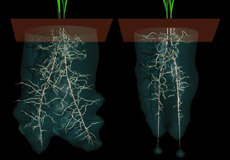
Modélisation de la plasticité environnementale. P. Prusinkiewicz, 1996, L-systèmes.
La croissance d’une plante peut être vue comme l’évolution d’un système dynamique dont l’état a une structure qui évolue elle-même dans le temps. Ces systèmes dynamiques à structure dynamique (SD)2 ont un comportement souvent complexe qui nécessite le développement de nouveaux outils d’analyse, de représentation et de simulation. Dans le cas de la modélisation des plantes, l’état d’une plante peut être représenté par une chaîne C de symboles où chaque symbole, M(x) se trouve dans un état x bien déterminé. La croissance de la plante se représente ainsi par l’allongement de cette chaîne de symboles. Pour représenter l’aspect ramifié des structures végétales, les chaînes de symboles considérées sont parenthésées, chaque niveau de parenthèse représentant un axe particulier de la plante.
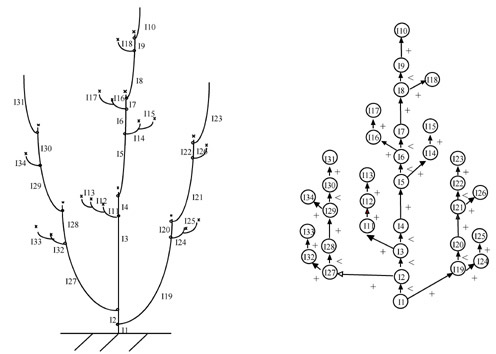
Ainsi, la structure topologique de la plante représentée ici peut être codée par la chaîne parenthésée suivante :
I1[I19[I24[I25]]I20[I21[I26]I22[I23]]]I2[I27[I32[I33]]I28[I29[I34]I30[I31]]]I3[I11[I12[I13]]] I4[I5[I14[I15]]I6[I16[I17]]I7[I8[I18]I9I10]]
L’évolution dans le temps de la plante peut donc être ramenée à la description de l’évolution d’une chaîne parenthésée. Ceci explique pourquoi les grammaires (ou systèmes de récriture) ont été largement utilisées pour modéliser la croissance ou l’organisation des végétaux. Le formalisme utilisé est celui des L-systèmes.
Le principe est le suivant :
On part d’un symbole initial M (appelé axiome) et on se donne des règles pour décrire les transformations de ce symbole à travers le temps. Par exemple, si M représente le méristème d’une plante, on peut écrire une règle de la forme :
M –> E M
qui spécifie qu’un méristème M produit, en un pas de temps, un « entrenœud » E suivi sur le même axe d’un nouveau méristème M. L’application successive de cette règle à un axiome initial M produit la séquence de résultats suivants :
M ==> E M ==> E E M ==> E E E M ==> E E E E M ==> E E E E E M ==> …
À chaque instant, l’organisme biologique (ici une plante) est représenté par une chaîne de symboles de cette séquence. L’état de l’organisme au pas de temps suivant est calculé en transformant simultanément tous les symboles de l’organisme à l’aide des règles spécifiées par la grammaire (dans cet exemple, les symboles E pour lesquels il n’y a pas de règle d’évolution explicitement définie sont considérés comme invariants à travers le temps). Ce processus permet de définir à chaque pas de temps un état valide de l’organisme.
L’état des différents constituants de la plante peut être représenté en attachant des variables aux différents symboles et en décrivant leur évolution dans l’écriture des règles. Par exemple :
M(t) –> E(1)M(t+1) E(d) –> E(d+1)
permet de décrire une plante dont le méristème apical vieillit à chaque pas de temps et dont le diamètre des entrenœuds grossit régulièrement :
M(0) ==> E(1)M(1) ==> E(2)E(1)M(2) ==> E(3)E(2)E(1)M(3) ==> E(4)E(3)E(2)E(1)M(4)==> …
L’application de certaines règles peut être contrôlée par des conditions portant sur l’état de la structure ou de son environnement :
M(t) :t<=3 –> EM(t+1)
M(t) :t>3 –> F
ce qui donne la séquence de développement suivante :
M(0) ==> EM(1) ==> EEM(2) ==> EEEM(3) ==> EEEEM(4) ==> EEEEF
Ici, le méristème apical se transforme au bout de 4 pas de temps en une fleur (symbole F), ce qui a pour effet d’arrêter le développement de la structure.
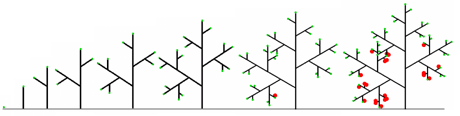
La possibilité de ramifier peut être rajoutée dans l’écriture de ces règles d’évolution, grâce à un parenthésage. Ainsi la règle :
M –> E[M]M
spécifie qu’un méristème M produit un entrenœud, un méristème axillaire et un nouveau méristème apical. Cette fois, l’application successive de cette règle à un axiome initial M produit la séquence de résultats suivants :
M ==> E[M]M ==> E[E[M]M]E[M]M ==> E[E[E[M]M]E[M]M]E[E[M]M]E[M]M ==> …
Chaque chaîne de cette séquence représente un stade du développement de la plante.
Interprétation géométrique des symboles.
En attachant une interprétation géométrique aux différents symboles d’une telle chaîne, il est possible de donner une représentation géométrique de la structure végétale à chaque instant du développement. Dans l’exemple suivant, chaque symbole pouvant constituer l’organisme, M, E, F est associé à une représentation géométrique. L’évolution de la plante dans le temps est décrite par deux règles :
M(t) :t < DUREE_DE_VIE –> E(t)[a(t)M(t+2)]M(t+1)
M(t) :t >= DUREE_DE_VIE –> F
Le méristème apical produit à chaque pas de temps un entrenœud et des méristèmes axillaires qui sont plus âgés que lui (a(t) représente l’angle phyllotaxique). Après une certaine période de temps, il se différencie en fleur (ce qui termine le développement de l’axe correspondant). Le résultat de l’application de ces règles à l’axiome M(0) est illustré sur la figure suivante.
Représentation géométrique de l’évolution de la structure végétale.
Cet exemple illustre un point très remarquable des systèmes dynamiques à structure dynamique. Il est parfois possible de décrire le développement de structures ramifiées relativement complexes avec très peu de règles.
Visionner la séquence vidéo (Vidéo Flash) Durée : 10 s © M. Renton
Le développement de ces concepts permet de modéliser de façon concise le développement de structures végétales beaucoup plus complexes, de tester des hypothèses de fonctionnement et d’analyser les propriétés de ces structures dynamiques, etc.
Et à cinq ans ?
Sur la base des projets en cours, nous pouvons prévoir en partie ce que sera devenue la technologie des plantes virtuelles à moyen terme (environ 5 ans). Nous saurons vraisemblablement par exemple :
- automatiser la mesure et la reconstruction informatique 3D des plantes (à partir de photographies ou de techniques d’observation au scanner laser) ;
- effectuer de premières évaluations des modèles de croissance sur des bases quantitatives ;
- passer aux premières applications agronomiques et forestières (par exemple concernant l’amélioration de la qualité du bois pour la construction, ou l’aide à la décision en agronomie) ;
- construire de nouveaux modèles biophysiques (interaction mécanique d’une plante avec le sol, étude de l’interaction des plantes avec le vent) ;
- mieux comprendre les mécanismes génétiques impliqués dans la croissance des plantes.
Dans 20 ans, imaginons…
Imaginons qu’avant d’acheter des graines ou des boutures chez le pépiniériste, nous puissions choisir ces graines sur un catalogue électronique et les planter artificiellement dans notre ordinateur. Imaginons que nous puissions ensuite suivre le développement de ces plantes sur l’ordinateur, tester différentes conditions de culture, observer les différents résultats paysagistes, agronomiques, forestiers ou environnementaux.
Imaginons qu’avant de croiser deux variétés, nous puissions savoir par simulation quel sera le (ou les) phénotype(s) de la nouvelle plante hybride. Imaginons que ceci nous permette de concevoir des plantes qui résistent à la sécheresse.
Imaginons simplement que nous comprenions les processus physiologiques à l’œuvre dans la croissance d’une plante, les chaînes biochimiques qui s’enchaînent pour contrôler le fonctionnement des cellules et des tissus, le rôle des gènes dans les processus morphogénétiques responsables de la différenciation cellulaire et de la création des différents organes. Imaginons que nous comprenions à travers quels mécanismes une telle activité à l’échelle moléculaire produit les structures observées à d’autres échelles (feuilles, branches, arbre), et aboutit à une certaine qualité de bois ou une certaine quantité de fruits.
Imaginons…
Remerciements
Les auteurs tiennent à remercier F. Boudon et C. Pradal pour avoir participé à l’élaboration de certaines illustrations ainsi que F. Sillion et C. Soler de l’INRIA Rhône-Alpes, Y. Caraglio et J.F. Barczi du CIRAD, A. Lacointe de l’INRA Clermont-Ferrand, M. Renton de l’Université de Queensland (Australie) et le Professeur Prusinkiewicz et son équipe de l’Université de Calgary, pour avoir mis à notre disposition différentes illustrations utilisées dans ce document.
Des articles scientifiques :
- Autran, D. et Traas, J. (2001) Organisation et fonctionnement des cellules souches végétales : le méristème apical d’Arabidopsis. Médecine/science 17, 836-844.
- Godin, C., Costes, E. et Sinoquet, H. (2005) Plant architecture modelling – virtual plants and complex systems. In: Turnbull, C. (ed.) Plant Architecture and its Manipulation. Blackwell, Annual Plant Reviews, Volume 17, sous presse.
- Prusinkiewicz, P., Hammel, M., Hanan, J. and Mech, R. (1997) Visual models of plant development. G. Rozenberg, A. Salomaa, eds. Springer-Verlag, Berlin, 535-598.
Des sites webs :
En France :
- UMR AMAP Botanique et Bioinformatique de l’Architecture des Plantes, CIRAD Montpellier
- UMR Biologie du développement des Espèces Pérennes Cultivées, INRA Montpellier
- UMR Physiologie Intégrée de l’Arbre Fruitier et Forestier, INRA Clermont-Ferrand
À l’étranger :
- Algorithmic Botany, University of Calgary
- Plant Architecture Informatics, University of Queensland
Niveau de lecture
Aidez-nous à évaluer le niveau de lecture de ce document.
Votre choix a été pris en compte. Merci d'avoir estimé le niveau de ce document !

Christophe Godin

Hervé Sinoquet
Directeur de l'UMR Physiologie Intégrée de l'Arbre Fruitier et Forestier (Clermont-Ferrand) jusqu'à son décès en septembre 2008.
Évelyne Costes
Chargée de Recherche INRA
Animatrice de l'équipe Architecture et Fonctionnement des Espèces Fruitières
UMR Biologie du développement des Espèces Pérennes Cultivées (Montpellier).
Jan Traas
Laboratoire de reproduction et développement de plantes, ENS (Lyon).